第三节 抗原的特异性
第三节 抗原的特异性
特异性(specificity)是指物质之间的相互吻合性、针对性和专一性。抗原的特异性表现在两个方面,即免疫原性的特异性和反应原性的特异性。前者是指某一特定抗原只能激发机体产生特定的免疫应答,即产生针对该抗原的特异性抗体和(或)致敏淋巴细胞;后者是指某一特定抗原只能与其相应的抗体和(或)致敏淋巴细胞特异性结合而出现反应。特异性是免疫应答最重要的特点,也是免疫学诊断与防治的理论依据。根据抗原、抗体反应具有特异性这一特点,可借助免疫学手段区分某些甚至用精细的化学方法都难以区别的物质之间的细微差异。决定抗原特异性的物质基础是抗原分子中的抗原决定簇。
一、抗原决定簇
抗原决定簇(antigenic determinant,AD)是指抗原分子中决定抗原特异性的特殊化学基团,又称表位(epitope)。抗原通过抗原决定簇与相应淋巴细胞表面的抗原受体结合,从而激活淋巴细胞,引起免疫应答;抗原也通过决定簇与相应抗体或致敏淋巴细胞发生特异性结合发挥免疫效应。因此,抗原决定簇是被免疫细胞识别的靶结构,也是免疫反应具有特异性的物质基础。抗原决定簇的性质、数目和空间构型决定着抗原的特异性。抗原分子表面决定簇的总数称为抗原结合价,又称功能价。大多数蛋白质抗原具有多个相同或不同的抗原决定簇,为多价抗原。一个多肽决定簇可由5~7个氨基酸残基组成,一种抗原决定簇只能刺激机体产生一种相应的抗体或致敏淋巴细胞。免疫应答反应与下列重要的抗原决定簇有直接关系。
-
T细胞和B细胞决定簇
在免疫应答中供T细胞抗原受体(TCR)识别的决定簇称T细胞决定簇。TCR只能识别10~20个氨基酸左右的多肽,此类决定簇一般不位于抗原分子表面,必须由抗原提呈细胞(APC)加工处理后才能被TCR识别。供B细胞抗原受体(BCR)或抗体识别的决定簇称B细胞决定簇。BCR或抗体能与未经APC加工处理的抗原发生反应,其识别的决定簇位于抗原分子表面,约由4~6个氨基酸残基或糖基组成。 -
载体决定簇与半抗原决定簇
天然抗原为含有蛋白质载体和大量半抗原的大分子。半抗原偶氮化后可结合到蛋白载体上,它具有载体和半抗原决定簇。每一种半抗原可理解为单一的抗原决定簇。半抗原只有结合到载体上,才能产生相应抗体,此现象称为载体效应,说明载体不是单纯起运载半抗原的作用,其本身也具有特异性。因此,一个完全抗原分子必须具有载体决定簇和半抗原决定簇。现已证明,在抗体形成过程中,T细胞识别载体决定簇,对抗体产生起辅助作用;B细胞识别半抗原决定簇,是产生抗体的细胞。
二、抗原-抗体反应的特异性
抗原-抗体反应的高度特异性可精确区分物质间极细微的差异。这种特异性不仅取决于抗原决定簇的化学组成,而且与化学基团的空间排列和构象密切相关。经人工抗原研究发现:
- 抗原-抗体反应犹如锁和钥匙,具有高度特异性:如由结合苯胺的抗原所激发机体产生的抗体,只能与苯胺决定簇起反应,而不能与其他抗原起反应。
- 特异性是由抗原决定簇所决定,而非由整个抗原分子决定:如将甲酸基、磺酸基、砷酸基连接在苯胺上,再经偶氮化与蛋白质结合后,将此化合物免疫动物得到的抗体只能与相应的基团起反应。
- 抗原决定簇的空间位置很重要,即使抗原决定簇相同,但其位置不同,特异性亦不同:如对位氨基苯甲酸蛋白抗原产生的抗体,只能与对位氨基苯甲酸蛋白抗原结合,而不能与邻位和间位的氨基苯甲酸蛋白抗原结合。
- 抗原结构的旋光度也与抗原特异性有关:如右旋酒石酸偶氮蛋白抗原激发产生的抗体,只能与右旋酒石酸偶氮蛋白抗原结合,而不能与左旋酒石酸偶氮蛋白抗原结合,这是因为二者的结合呈互补方式,犹如左手的手套不能适合于右手一样。
天然单纯蛋白质抗原,其抗原特异性主要取决于末端氨基酸序列的不同。不同种属动物血清白蛋白因其末端氨基酸序列不同,即表现出种属特异性的差异。
三、共同抗原与交叉反应
天然抗原表面常带有多种抗原决定簇,每种决定簇都能刺激机体产生一种特异性抗体,因此复杂抗原能使机体产生多种特异性抗体。如伤寒患者血清中可检出针对伤寒杆菌鞭毛的抗体、抗表面成分的抗体及多种菌体抗体。不同抗原相互间可存在部分相同的抗原决定簇,称为共同抗原。亲缘关系很近的生物间存在的共同抗原,称为类属抗原;无种属关系的生物间存在的共同抗原称为异嗜性抗原。
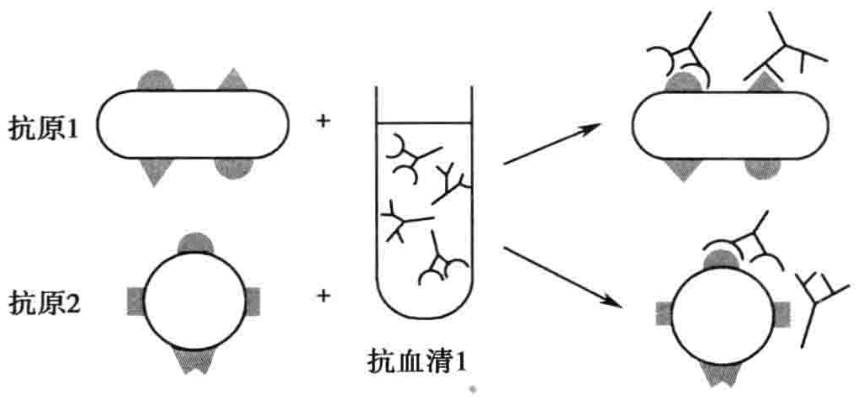
抗原(或抗体)除与其相应抗体(或抗原)发生特异性反应外,有时还可与其他抗体(或抗原)发生反应,称为交叉反应(图2-1)。交叉反应不仅在两种抗原决定簇构型完全相同时发生,也可在两种决定簇构型相似的情况下发生,只是后者的结合力相对较弱。血清学诊断中出现交叉反应时,易造成判断上的混乱,给免疫学诊断带来困难。但根据交叉反应的原理,利用容易得到的某种共同抗原(或抗体),在血清学诊断中检测体内相应的抗体(或抗原),在临床疾病的辅助诊断上有重要作用。