第二节 MHC编码的分子
第二节 MHC 编码的分子
由 MHC 编码的蛋白分子称为 MHC 分子或 MHC 抗原,在人类也称为人类白细胞抗原(HLA)。借助于 X 线晶体衍射技术,对 MHC 分子立体结构的认识已经很清楚,其主要生物学功能也相对明确。
一、MHC 分子的分布
MHC I 类分子广泛分布于体内所有有核细胞表面,包括网织红细胞和血小板。成熟的红细胞一般不表达 HLA 抗原。淋巴细胞表面 MHC 分子的密度最大。
MHC II 类分子主要分布于抗原提呈细胞(如树突状细胞、单核-巨噬细胞、B 细胞)和活化的 T 细胞的表面。
MHC III 类分子主要是一些存在于血清及其他体液中的可溶性分子,包括某些补体成分(如 C2、C4、Bf)、TNF、LT 以及 HSP 等。
二、MHC 分子的结构
1. MHC I 类分子的结构 MHC I 类分子是由 α 链(重链)和 β 链(轻链)通过非共价键连接组成的异二聚体。α 链胞外区从 N 端起依次由 α1、α2 和 α3 结构域组成。β 链为 β2 微球蛋白(β2-microglobulin,β2m),仅有一个结构域。α1 和 α2 构成 MHC I 类分子的抗原肽结合区(又称肽结合槽),通过 α3 的延伸部分锚定于细胞膜。α3 结构域序列高度保守,是 I 类分子与 T 细胞表面 CD8 分子相互结合的结构域。I 类分子 α 链羧基末端含有约 30 个氨基酸残基,位于胞质中,高度保守,可能参与调解 I 类分子与其他膜蛋白或细胞骨架成分间的相互作用,也与细胞跨膜信号传递有关。β2 微球蛋白无跨膜区,仅以非共价键与 α3 结构域相互作用,对维持 I 类分子天然构型的稳定性具有重要作用。
2. MHC II 类分子的结构 MHC II 类分子是由 α 链(重链)和 β 链(轻链)通过非共价结合组成的异二聚体。MHC II 类分子的胞外区由 α1、α2、β1 和 β2 四个结构域组成。α1 和 β1 构成 MHC II 类分子的抗原肽结合区,通过 α2 和 β2 的延伸部分锚定于细胞膜,并通过 β2 与 T 细胞表面 CD4 相互作用。与 I 类分子相比,II 类分子抗原肽结合区两端更为开放,可以容纳更多的氨基酸残基。MHC II 类分子羧基端位于胞质中,含有 10~15 个氨基酸残基,可能参与细胞跨膜信号传递。HLA I 类分子和 II 类分子的结构如图 5-2 所示。
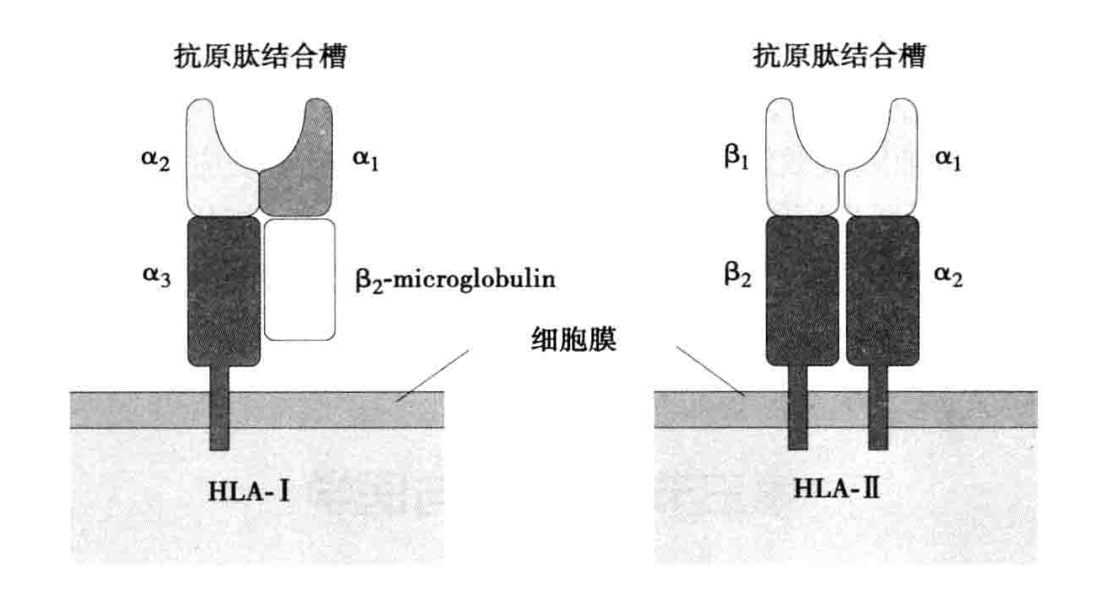
人类 HLA I 类分子与 II 类分子的结构特征比较(表 5-1)。
| 特征 | HLA I 类分子 | HLA II 类分子 |
|---|---|---|
| 多肽链 | α 链,β2m | α 链,β 链 |
| 多态性位点 | α1,α2 结构域 | α1,β1 结构域 |
| 与 T 细胞结合位点 | CD8 与 α3 结合 | CD4 与 β2 结合 |
| 结合的抗原肽 | 8~11 氨基酸残基 | 10~30 氨基酸残基 |
| 类别 | HLA-A,HLA-B,HLA-C | HLA-DR,HLA-DQ,HLA-DP |
| 组织分布 | 所有有核细胞表面 | DC、MΦ、B 细胞等 APC 以及活化的 T 细胞表面 |
| 功能 | 提呈内源性抗原,激发 CD8⁺T 细胞应答 | 提呈外源性抗原,激发 CD4⁺T 细胞应答 |
三、MHC 分子的免疫功能
1. 提呈抗原 提呈抗原是 MHC 分子的最主要功能之一。内源性抗原/外源性抗原首先由抗原提呈细胞(APC)加工为小分子抗原肽,后者结合于 MHC I 类/II 类分子的抗原结合槽,形成抗原肽-MHC I 类/II 类分子复合物(pMHC),然后被转运并表达于 APC 表面,分别供 CD8+ T 细胞/CD4+ T 细胞识别,进而启动适应性免疫应答(详见第六章)。通常情况下,T 细胞仅能识别 APC 表面提呈的 pMHC。
2. 参与 T 细胞分化、发育 进入胸腺的淋巴样干细胞在胸腺微环境作用下逐步发育、分化为成熟 T 细胞,MHC 分子通过介导胸腺细胞的阳性选择与阴性选择,使 T 细胞最终分化为 CD4+ T 细胞和 CD8+ T 细胞,并获得 MHC 限制性和对自身抗原的中枢性耐受。
3. 作为调节分子参与固有免疫应答
(1)参与补体反应和炎症反应 MHC III 类基因作为部分补体成分的编码基因,参与补体反应和免疫性疾病的调控。炎症相关基因参与启动和调控炎症反应,并在应激反应中发挥作用。
(2)参与调节 NK 细胞活性 表达于自身细胞表面的 MHC I 类分子可结合 NK 细胞表面的抑制性受体(KIR),从而启动抑制性信号,抑制 NK 细胞的杀伤活性,故正常组织细胞不会受到 NK 细胞攻击;而某些靶细胞(肿瘤细胞、病毒感染细胞、移植物组织细胞等)表面的 MHC I 类分子表达减少或缺失,则 NK 细胞的抑制性信号被阻断,导致 NK 细胞被激活,从而杀伤靶细胞。
(3)参与免疫应答的遗传调控 不同个体对特定抗原物质是否产生应答以及应答的强弱存在差异,这与 MHC 的高度多态性相关。MHC 高度多态性导致其编码的 MHC 分子氨基酸序列、蛋白结构多变,进而导致群体中不同个体对抗原的结合能力出现差异,由此实现 MHC 在群体水平对免疫应答的遗传调控,也有利于推动生命的进化。