第二节 体液免疫应答
第二节 体液免疫应答
B 细胞接受抗原刺激后转化为浆细胞分泌抗体,进而产生免疫效应,因抗体存在于血清等各种体液中,因此由浆细胞分泌抗体介导的特异性免疫应答称为体液免疫应答。体液免疫应答可因抗原类型不同,而有不同的免疫应答特征。
一、TD 抗原诱导的体液免疫应答
TD 抗原引起的体液免疫应答至少需要四种免疫细胞,即树突状细胞、CD4+ 初始 T 细胞、Th2 细胞和 B 细胞参与。
(一)感应阶段
抗原初次进人机体一般由树突状细胞(DC)摄取、加工处理后以抗原肽-MHC 分子复合体(pMHC)形式提呈给 CD4+ Th 细胞。抗原再次进人机体则主要由单核-巨噬细胞或 B 细胞提呈给 CD4+ Th 细胞。B 细胞可通过 BCR 直接识别抗原决定簇,获取抗原信息。
(二)反应阶段
B 细胞在 CD4+ Th 细胞辅助下,通过双信号模式增殖分化为浆细胞,并在不同细胞因子的作用下产生不同类型的抗体。其中有一部分 B 细胞成为记忆细胞(Bm)。当相同抗原再次刺激时,Bm 可迅速增殖分化为浆细胞,产生再次免疫应答。
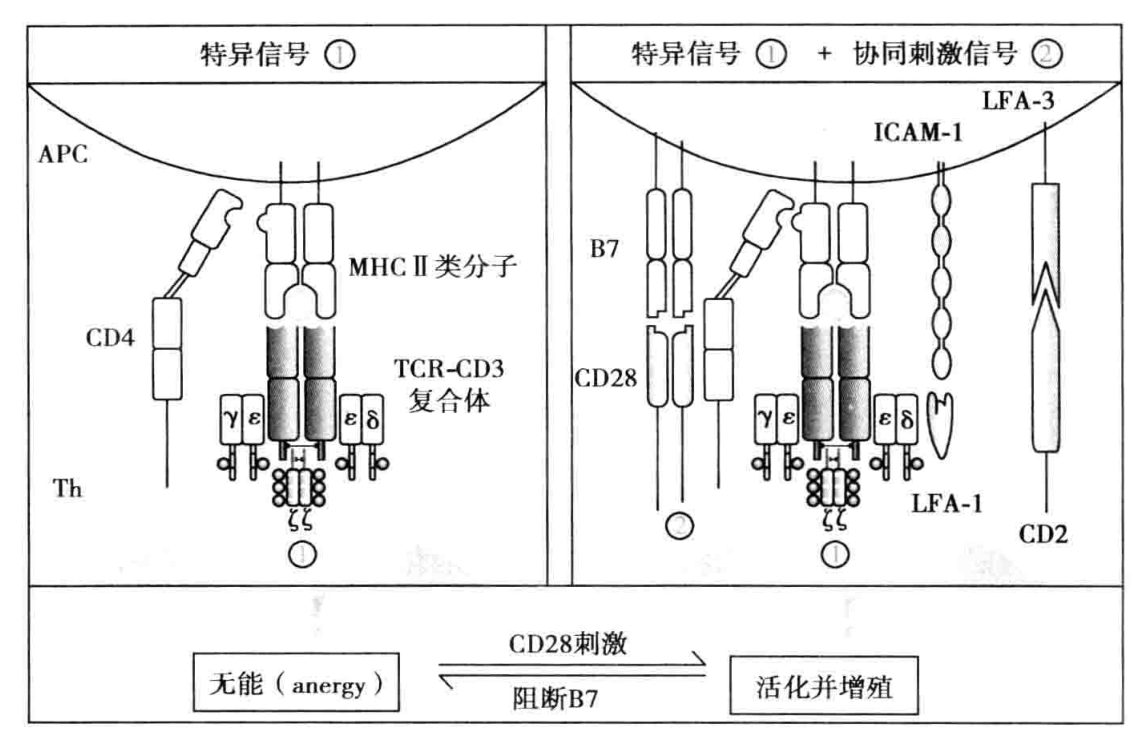
- CD4+ Th 细胞活化、增殖与分化 CD4+ Th 细胞的完全活化需要双信号和某些细胞因子的作用。 (1)第一活化信号:即抗原识别信号。CD4+ Th 细胞通过 TCR 识别 APC 表面 pMHC 中的抗原肽,同时 CD4 分子识别 APC 表面的 MHC II 类分子,获得第一活化信号(双识别)。 (2)第二活化信号:即协同刺激信号。CD4+ Th 通过表面 CD28 和 LFA-1 等共刺激分子与 B 细胞表面相应 B7 和 ICAM-1 等共刺激分子结合,可诱导产生 T 细胞活化第二信号使 CD4+ Th 细胞激活。活化 CD4+ Th 细胞可表达 CD40L 和多种细胞因子的受体,同时分泌 IL-2、IL-4、IL-5 和 IFN-γ 等多种细胞因子发挥免疫调节作用。只有抗原识别信号而缺乏协同刺激信号的 Th 细胞不能被活化,导致克隆无能(图 6-1)。 (3)细胞因子的作用:除双信号外,由活化的 APC 和 T 细胞产生的 IL-1、IL-2、IL-6、IL-12 等细胞因子有利于 Th 细胞充分活化。
-
B 细胞活化、增殖与分化 在活化 CD4+ Th 细胞的辅助下,B 细胞可通过双信号模式并在相应细胞因子的作用下完全活化。 (1)第一活化信号:即抗原识别信号。B 细胞可通过 BCR 直接识别天然抗原表位,产生第一活化信号。 (2)第二活化信号:即协同刺激信号。B 细胞通过表面 CD40 和 ICAM-1 等共刺激分子与活化 CD4+ Th 细胞表面相应的 CD40L 和 LFA-1 等共刺激分子结合,可诱导产生 B 细胞活化第二信号使 B 细胞活化。活化 B 细胞可表达多种细胞因子受体,促其进一步增殖分化。 (3)细胞因子的作用:Th 细胞产生的多种细胞因子如 IL-2、IL-4、IL-5、IL-6 等有助于 B 增殖、分化为浆细胞。其中部分 B 细胞分化为记忆 B 细胞(Bm)。
-
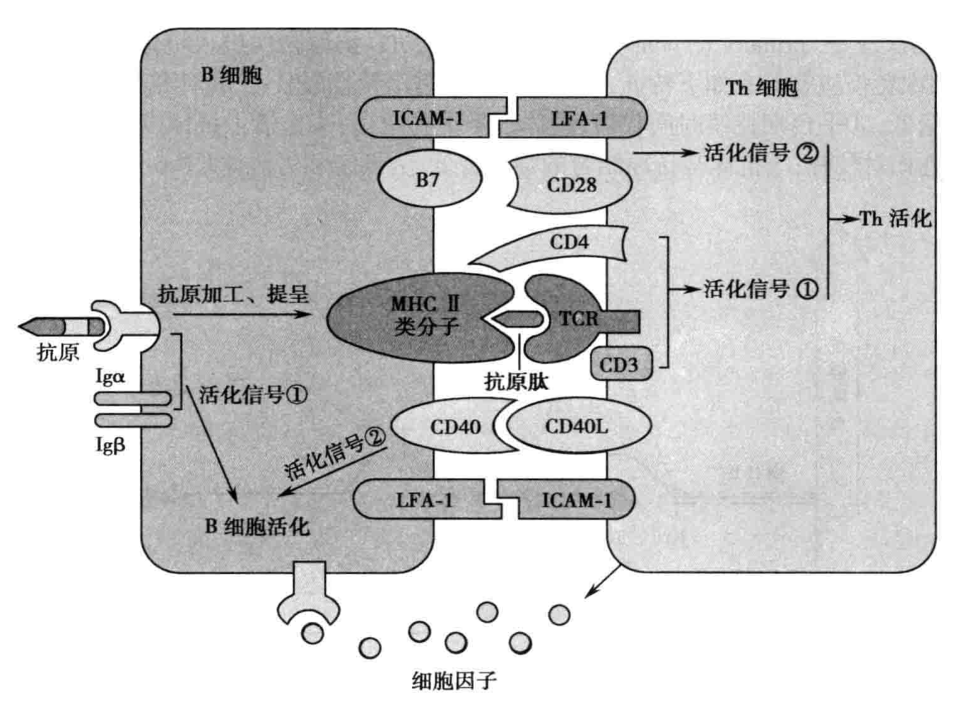
Th 细胞与 B 细胞间相互作用 在反应阶段,B 细胞与 Th 细胞通过细胞间膜分子接触及分泌细胞因子相互作用(图 6-2)。 (1)B 细胞对 Th 细胞的作用:B 细胞作为专职 APC,提呈抗原肽供 CD4+ Th 细胞识别,Th 细胞通过双识别获得第一活化信号;B 细胞表面 B7 和 ICAM-1 等共刺激分子与 Th 细胞表面相应 CD28 和 LFA-1 等共刺激分子结合,可诱导产生 T 细胞活化第二信号使 CD4+ Th 细胞激活。 (2)Th 细胞对 B 细胞的作用:活化的 CD4+ Th 细胞表面表达 CD40L 和 LFA-1 等共刺激分子,与 B 细胞表面 CD40 和 ICAM-1 等共刺激分子结合,使 B 细胞获得第二活化信号。此外,活化 CD4+ Th 细胞可分泌 IL-2、IL-4、IL-5 和 IL-6 等多种细胞因子,促进 B 细胞活化、增殖、分化。
(三)效应阶段
此阶段是抗体发挥生物学效应的阶段。浆细胞合成分泌抗体后,与相应抗原结合发挥中和、调理吞噬、激活补体、ADCC 等多种免疫效应,最终清除抗原异物。
二、TI 抗原诱导的体液免疫应答
TI 抗原可直接作用于 B 细胞产生体液免疫应答,而无需 Th 细胞和抗原提呈细胞(APC)参与。根据抗原分子结构不同,TI 抗原可分为 TI-1 抗原(TI-1Ag)和 TI-2 抗原(TI-2Ag),它们分别以不同机制激活 B 细胞。
- TI-1Ag 激活 B 细胞 细菌 LPS 和多聚鞭毛等多为 TI-1Ag。此类抗原具有特异性抗原表位和 B 细胞丝裂原两种不同的抗原结构。TI-1Ag 激活 B 细胞需要双信号:① B 细胞表面 BCR 识别结合 TI-1Ag 特异性抗原表位,产生第一活化信号;② B 细胞表面丝裂原受体结合 TI-1Ag 相应的丝裂原,产生第二活化信号激活 B 细胞。
- TI-2Ag 激活 B 细胞 细菌细胞壁和荚膜多糖等多为 TI-2Ag。此类抗原只具有高密度重复排列的相同抗原决定簇,无 B 细胞丝裂原。TI-2Ag 上多个相同的抗原决定簇与 B 细胞表面 BCR 广泛交联结合,可直接诱导 B 细胞活化。
TI-Ag 诱导 B 细胞产生的体液免疫应答的特点包括:① 不需要抗原提呈细胞的提呈;② 不需要 Th 细胞的辅助;③ 不产生记忆细胞,无再次应答效应;④ 只产生 IgM 类别的抗体。
三、抗体产生的一般规律
抗体产生过程可分为四个阶段:
① 潜伏期:指抗原进人体内到相应抗体产生之前的阶段,此期的长短与抗原的性质、抗原进人途径、佐剂类型和机体免疫状态等因素有关,短者几天,长可数周; ② 对数期:指抗体呈指数增长的阶段; ③ 平台期:指抗体水平相对稳定,既不明显增高,也不明显减少的阶段; ④ 下降期:指抗体合成率小于降解速度,血清中抗体水平逐渐下降的阶段,此期可持续几天或几周。
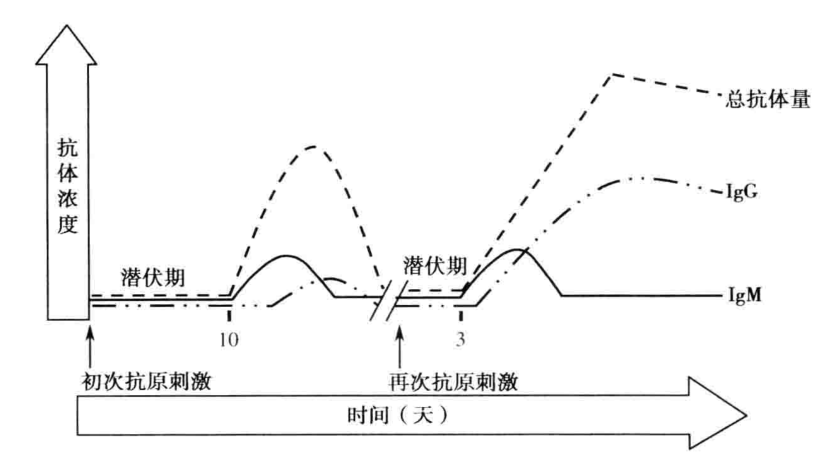
- 初次应答(primary response) 指病原生物等 TD 抗原初次进人机体诱发的体液免疫应答。初次免疫应答具有如下特征:① 抗体产生所需潜伏期较长;② 抗体倍增所需时间较长,抗体含量低;③ 平台期持续时间较短,抗体水平下降迅速;④ 血清中抗体以 IgM 为主,IgG 为辅且出现相对较晚;⑤ 抗体与抗原结合的强度较低,为低亲和力抗体(图 6-3)。
- 再次应答(secondary immune response) 指初次应答后,机体再次接受相同抗原刺激产生的体液免疫应答。与初次应答相比,再次应答具有如下特征:① 潜伏期明显缩短,约为初次应答潜伏期时间的一半;② 抗体倍增所需时间短,抗体含量迅速大幅度上升;③ 平台期抗体浓度高且维持时间较长,抗体水平下降缓慢;④ 用较少量抗原刺激即可诱发再次应答;⑤ 血清中抗体以 IgG 为主;⑥ 抗体与抗原结合的强度较高,为高亲和力抗体且较均一(表 6-1)。
| 特点 | 初次应答 | 再次应答 |
|---|---|---|
| 潜伏期 | 长,1-2 周 | 短,2-3 天 |
| 抗体类别 | 以 IgM 类为主 | 以 IgG 类为主 |
| 抗体滴度 | 低 | 高 |
| 抗体亲和力 | 低 | 高 |
| 抗体维持时间 | 短 | 长 |
表6-1 初次应答与再次应答抗体产生规律的比较
- 抗体产生规律的临床意义 再次应答主要由 Tm、Bm 介导产生,其应答规律已广泛应用于医学实践:① 在免疫血清的制备和疫苗接种中,常通过再次或多次加强免疫,诱导高效价、高亲和力的抗体,增强免疫效果;② 在某些感染性疾病的免疫学诊断中,血清 IgM 抗体升高可作为早期感染诊断的依据之一;③ 检测血清抗体含量变化有助于了解病程与疾病转归,IgG 类抗体或总抗体效价增高 4 倍以上时具有诊断意义。
四、体液免疫的生物学效应
体液免疫应答的主要效应分子为抗体,抗体可发挥多种生物学效应:
① 中和作用:抗体可直接中和外毒素或病毒; ② 激活补体系统:抗体与抗原结合形成的免疫复合物,可激活补体系统,溶解靶细胞; ③ 调理作用:增强吞噬细胞的吞噬与杀伤功能; ④ 参与 ADCC 效应:增强 NK 细胞、Mφ 等细胞对靶细胞的杀伤作用; ⑤ 参与超敏反应:IgE、IgM、IgG、IgA 等抗体可参与 I、II、III 型超敏反应,引起组织病理损伤; ⑥ 参与黏膜局部免疫:IgA 可在黏膜局部发挥免疫保护作用。